|
Cytokinins have been found
in almost all higher plants as well as mosses, fungi, bacteria,
and also in tRNA of many prokaryotes and eukaryotes. Today there
are more than 200 natural and synthetic cytokinins combined. Cytokinin
concentrations are highest in meristematic regions and areas of
continuous growth potential such as roots, young leaves, developing
fruits, and seeds (Arteca, 1996; Mauseth, 1991; Raven, 1992; Salisbury
and Ross, 1992).
History of Cytokinins
In 1913, Gottlieb Haberlandt discovered that
a compound found in phloem had the ability to stimulate cell division
(Haberlandt, 1913). In 1941, Johannes van Overbeek discovered that
the milky endosperm from coconut also had this ability. He also
showed that various other plant species had compounds which stimulated
cell division (van Overbeek, 1941). In 1954, Jablonski and Skoog
extended the work of Haberlandt showing that vascular tissues contained
compounds which promote cell division (Jablonski and Skoog, 1954).
The first cytokinin was isolated from herring sperm in 1955 by Miller
and his associates (Miller et al., 1955). This compound was named
kinetin because of its ability to promote cytokinesis. Hall and
deRopp reported that kinetin could be formed from DNA degradation
products in 1955 (Hall and deRopp, 1955). The first naturally occurring
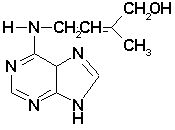
cytokinin was isolated from corn in 1961 by Miller (Miller, 1961).
It was later called zeatin. Almost simultaneous with Miller Letham
published a report on zeatin as a factor inducing cell division
and later described its chemical properties (Letham, 1963). It is
Miller and Letham that are credited with the simultaneous discovery
of zeatin. Since that time, many more naturally occurring cytokinins
have been isolated and the compound is ubiquitous to all plant species
in one form or another (Arteca, 1996; Salisbury and Ross, 1992).
Biosynthesis and Metabolism of Cytokinins
Cytokinin is generally found in higher concentrations
in meristematic regions and growing tissues. They are believed to
be synthesized in the roots and translocated via the xylem to shoots.
Cytokinin biosynthesis happens through the biochemical modification
of adenine. The process by which they are synthesized is as follows
(McGaw, 1995; Salisbury and Ross, 1992):
A product of the mevalonate pathway called isopentyl pyrophosphate
is isomerized.
This isomer can then react with adenosine monophosphate with the
aid of an enzyme called isopentenyl AMP synthase.
The result is isopentenyl adenosine-5'-phosphate (isopentenyl AMP).
This product can then be converted to isopentenyl adenosine by removal
of the phosphate by a phosphatase and further converted to isopentenyl
adenine by removal of the ribose group.
Isopentenyl adenine can be converted to the three major forms of
naturally occurring cytokinins.
Other pathways or slight alterations of this one probably lead to
the other forms.
Degradation of cytokinins occurs largely due to the enzyme cytokinin
oxidase. This enzyme removes the side chain and releases adenine.
Derivitives can also be made but the pathways are more complex and
poorly understood.
Cytokinin Functions
A list of some of the known physiological
effects caused by cytokinins are listed below. The response will
vary depending on the type of cytokinin and plant species (Davies,
1995; Mauseth, 1991; Raven, 1992; Salisbury and Ross, 1992).
- Stimulates cell division.
- Stimulates morphogenesis
(shoot initiation/bud formation) in tissue culture.
- Stimulates the growth
of lateral buds-release of apical dominance.
- Stimulates leaf expansion
resulting from cell enlargement.
- May enhance stomatal
opening in some species.
- Promotes the conversion
of etioplasts into chloroplasts via stimulation of chlorophyll
synthesis.
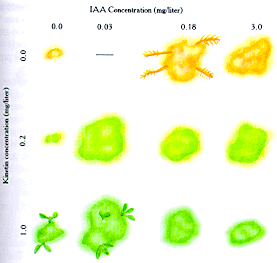
The illustration above shows the effect of cytokinin and auxin concentration
on tissue culture experiments (Mauseth, 1991)
|